Pollen grains and spores form the basis of another important aspect of palaeoclimate reconstruction, generally referred to as pollen analysis, or palynology. Where pollen and spores have accumulated over time, a record of the past vegetation of an area may be preserved. Often, changes in the vegetation of an area may be due to changes of climate. Interpreting past vegetation through pollen analysis may therefore offer a form of palaeoclimatic reconstruction.
Pollen grains and spores are extremely resistant to decay and are produced in huge quantities which are distributed widely from their source. A particular genus or species of plant may possess unique morphological characteristics to aid the reconstruction of past vegetation assemblages.
Differences in pollen productivity and dispersion rates pose a significant problem for palaeoclimatic reconstruction because the relative abundances of pollen grains in a deposit cannot be directly interpreted in terms of species abundance in the study area. For example, a vegetation community composed of 15% x, 35% y and 50 % z may be represented in a deposit by approximately equal amounts of pollen from x, y and z, because of the inter-species differences in pollen productivity and dispersion. To resolve this, it is necessary to have some function that relates pollen abundance and spatial distribution to species frequency. The various approaches to this have employed the uniformitarian principle i.e. the “present is the key to the past” (e.g. Davis, 1963), although the evidence that this assumption is valid is by no means conclusive.
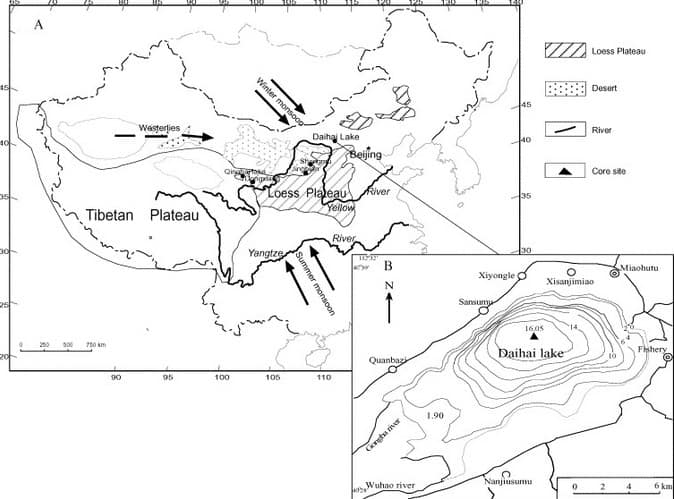
Pollen being an aeolian (wind-blown) sediment will accumulate on any undisturbed surface. Sediments containing fossil pollen have been taken from peat bogs, lake beds, alluvial deposits, ocean bottoms and ice cores (Jacobson & Bradshaw, 1981). Where pollen has been deposited in water, one must be aware of the non-climatic effects that cause variations in pollen type and abundances. These include differential settling, turbulence mixing and the effects of burrowing organisms (Davis, 1973).
Unfortunately, the difficulties associated with pollen analysis have meant that most palaeoclimate reconstructions have proceeded in a qualitative way only – the climate was wetter/drier or warmer/colder. Sometimes it may be possible to quantify palaeoclimatic variations by the use, not of total pollen assemblages, but of individual indicator species, plants which may not be abundant but which are thought to be limited by specific climatic conditions (e.g. holly (Ilex), ivy (Hedera) and mistletoe (Viscum) (Iversen, 1944)).


Leave a Reply